DINÁMICA DE POBLACIONES
1. Fenología de las plagas
El estudio de la dinámica de
poblaciones ocupa la parte central de todo proyecto de MIP, ya que al depender
del control natural, la dinámica define los umbrales de daño y económico. La
fenología de un organismo es todo lo que le sucede durante su ciclo de vida,
frente al medio que le rodea, sea de carácter biótico o abiótico. Así, la
fenología de una plaga es el recuento de todo lo que le sucede, como individuo
o población, con relación a su hospedante, parasitoides, depredadores,
patógenos, heladas, lluvias y el resto de los fenómenos naturales o inducidos,
como la tecnología agrícola.
Los aspectos que debemos conocer
de una población son: su taxonomía, biología, ecología y comportamiento
–etología, tanto a nivel autoecológico como sinecológico. Esto nos permitirá
identificar los detalles relevantes en la vida del individuo como unidad mínima
del sistema o subsistema en que se ubica, o en la población.
1.1. Taxonomía
El estudio de una plaga debe
comenzar con la determinación de su especie, y para los fines del MIP, la
especie de todos los organismos
coevolucionados, sean hospedantes, competidores, parasitoides,
depredadores o patógenos.
1.2. Ciclos biológicos
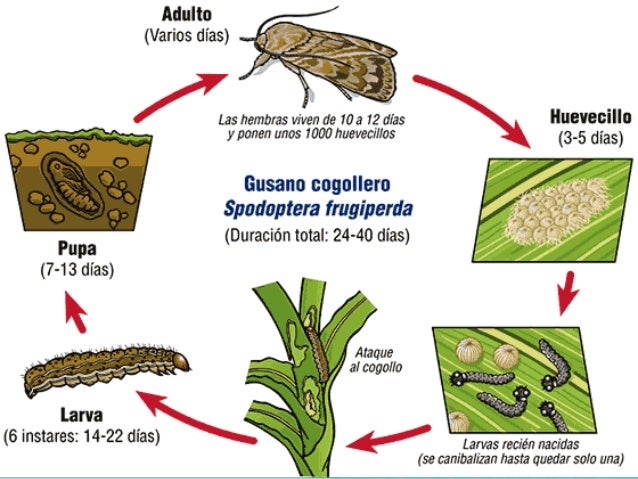
El éxito reproductivo de las especies depende de las tasas de nacimiento, reproducción y muerte. En los insectos el ciclo biológico varía desde especies en casi continua reproducción durante periodos muy cortos de vida (pulgones en partenogénesis), hasta especies de reproducción única en varios lustros (cigarras periódicas).
Se identifican tres
características que influyen en el número de descendientes de una sola hembra:
·
Número de descendientes por evento reproductivo.
·
Número de eventos reproductivos durante el ciclo
biológico (monovoltinismo a multivoltinismo).
·
La edad en que ocurre la primera reproducción.
Los insectos que tienden a tener
muchos descendientes por evento, muchos eventos (generaciones) y hacerlo a temprana edad, son más oportunistas (estrategas r o de ciclo biológico oportunista).
Mientras que aquellos que tienden a tener pocos descendientes, pocos eventos
reproductivos, y a reproducirse lo más tarde posible, se les llama equilibristas (estrategas k o de ciclo biológico equilibrista.
Las plagas de cultivos
estacionales y anuales son oportunistas, y empiezan con una infestación inicial
que siempre es “extranjera” (aloinfestación). Las de los cultivos perennes son equilibristas
(estratega k), tienen una tasa reproductiva menor, y "nunca"
abandonan el hospedante manteniendo “constantes” sus poblaciones
autoinfestantes; sólo los cunden cuando la estación les es muy favorable. Estos
cultivos pueden ser aloinfestados en las partes que se renuevan estacionalmente.
El rasgo más sobresaliente del
ciclo biológico es la meiosis y, por lo tanto, la forma sexual de reproducción,
presente en la mayoría de los insectos. La reproducción asexual, a pesar de ser
mucho más simple, no fue elegida por la evolución como la estrategia más común
de reproducción, obviamente debido a que reduce las posibilidades de
“supervivencia del más apto”.
1.3. Ecología poblacional
Se refiere al estudio de la interacción de los organismos con su ambiente, a su distribución y su abundancia.
El estudio de la
ecología de las plagas deberá responder a las siguientes preguntas:
¿Cuáles son las interacciones del individuo con su
ambiente biótico y abiótico?, ¿Cuáles las de la población?
¿Cómo modifican su ambiente?; ¿cómo los modifica
él?
¿Qué factores ambientales determinan la presencia
de tal o cuál especie?
¿Cuáles determinan sus cantidades?
Las respuestas a esas
interrogantes son la médula de los estudios ecológicos de las plagas, y deben
comenzar a gestarse en el ámbito individual. La autoecología (o ecología
fisiológica, o ecología del organismo, o ecología organísmica) de una plaga se
centra en el estudio de las maneras en que el individuo responde morfológica, fisiológica
y conductualmente a los retos que le impone su ambiente físico-químico−biológico−tecnológico.
Y en el de los límites individuales de tolerancia a cada una de las diferentes
presiones de selección, representadas por factores:
- físicos y químicos (humedad, temperatura, luminosidad, pH, etc.);
- meteorológicos (heladas, sequías, granizo, vientos, etc.),
- de competencia inter e intraespecífica;
- de parasitoidismo y depredación;
- de parasitismo por patógenos;
- de calidad alimentaria (consumo) de su hospedante;
- tecnológicos (cómo afecta al individuo la tecnología agrícola: desde la forma de cultivar, hasta el control con cualquier táctica).
Conocida la autoecología del
individuo, se pasa a la ecología de su población, y al análisis de sus límites
de tolerancia a las mismas presiones de selección, así como al análisis de las comunidades
(organismos) con quienes interacciona. A este nivel, las interrogantes se
refieren a cómo los actos de depredación, parasitismo, competencia y otros
tipos de interacción, modifican la estructura y organización de las poblaciones.
1.4. Comportamiento
¿Cómo podríamos saber la adaptabilidad o adaptación de una plaga a su hospedante y su agroecosistema, sin estudiarlos profundamente?
La técnica del insecto estéril,
aplicada a la “mosca del mediterráneo” Ceratitis
capitata y al“gusano de las heridas del ganado” Cochliomya hominivorax son magníficos ejemplos de lo que una sola
táctica de combate (control genético autocida) puede hacer contra una plaga, cuando
se basa en el conocimiento profundo de su comportamiento.
El comportamiento es la
manifestación de máxima adaptabilidad genética, de origen evolutivo, para
alimentarse, reproducirse y sobrevivir. Siendo genético y evolutivo está sujeto
a la selección natural, incluso cuando es aprendido en un contexto ecológico
específico. Por lo tanto debemos estudiar ecología conductual (el
comportamiento de origen evolutivo-ecológico) si es que tarde o temprano vamos
a hacer MIP. Un aspecto importante en el estudio del comportamiento es ecología
química de la plaga, en relación a sus conespecíficos, así como a su
hospedante, parasitoides y depredadores. Nos referimos, a los semioquímicos
involucrados: sinomonas, cairomonas; feromonas, alomonas y demás.
2. Localización y temporalidad de las plagas
Las poblaciones ocupan un lugar físico determinado (localización) en el momento de muestrearlas (temporalidad), y tienen un tamaño numérico. Al referir las poblaciones calculadas a la superficie o sustrato en que se hallaron, tenemos un cálculo de su densidad. Ambos parámetros, tamaño y densidad son determinables por muestreo. Refirámonos a la localización espacio-temporal de las plagas.
2.1. Localización
Se refiere a dónde buscar a la
plaga que se va a muestrear y cuya densidad va a medirse. Cuál es su localización
en la planta o ambiente; dónde se alimenta, refugia, copula, o hiberna. Generalmente
el estadio más perjudicial es un inmaduro (ninfa y larva solitarias o agregadas
que se localiza, con excepciones, en el órgano afectado). Los adultos,
individuales o en “colonia”, pueden ocupar estratos diferentes de los que
ocuparon sus inmaduros en la misma planta. En los casos de insectos de
metamorfosis completa puede haber un tercer estrato, el de pupación. El
panorama se complica algo más cuando los insectos son migratorios, y todavía
más cuando, además de migrar, entran en diapausa o invernan (a veces más de
medio año). En todo caso la estructura de edades (población relativa de cada
instar), más las constantes de muerte, nacimiento y crecimiento poblacional,
forman la demografía de la especie, misma que debe ser considerada por
localidades (sin olvidar la proporción de sexos).
2.2. Temporalidad de las plagas
En el tiempo, los artrópodos plaga se distribuyen aproximadamente como su hospedante, asumiendo dos tipos generales de estrategias:
La estrategia r, más modernamente llamada coyuntural,
que caracteriza a los individuos que se reproducen por estación (en hospedantes
efímeros), o durante el lapso que un hospedante perenne tiene renuevos. Y lo
hacen en grandes cantidades, a bajo costo biológico, con períodos cortos de vida
y asumiendo tamaño individual pequeño. En este caso son frecuentes las
explosiones poblacionales, dependiendo del tipo de cultivo, y dependiendo del
voltinismo o número de generaciones de la plaga “por temporada”. Generalmente predomina
el multivoltinismo (varias generaciones en la estación o el lapso), pero puede
haber bivoltinismo y, ocasionalmente, puede haber estrategas r que son mono o
univoltinos (una generación anual, donde el insecto puede pasar gran parte del
ciclo en estadio diapáusico adulto, como sucede en el altiplano con la
conchuela del frijol Epilachna varivestis).
La estrategia k caracteriza a los organismos que
aproximadamente están en equilibrio con el hospedante, razón por la que,
modernamente, se les llama “equilibristas”. Estos no alcanzan a formar grandes
poblaciones; “nunca” abandonan al hospedante; y son relativamente grandes de tamaño.
En este caso el voltinismo funciona al revés: predomina el univoltinismo, puede
haber bivoltinismo y es raro el voltinismo múltiple. Excepciones notables de
voltinismo son los insectos que tienen una generación cada dos o más años,
sobresaliendo el caso de las cigarras periódicas. El ciclo más corto conocido
de una cigarra periódica es de cuatro años; algunas llegan a 17.
La diferencia fundamental
estribaría en que el r interrumpe su relación con el hospedante y el k la
mantiene constante; en otras palabras, el primero es epidémico y el segundo,
asumiendo una relación permanente con el
hospedante, es endémico.
La distribución temporal, por lo
visto, depende de los ciclos temporales del hospedante y de la adaptación de
sus plagas a él. Es obvio que la distribución de los depredadores y
parasitoides sigue patrones semejantes, según su especificidad. Todas las
plagas oportunistas pasan parte de su ciclo fuera del hospedante, es decir, tienen que aloinfestarlo
inicialmente. Las equilibristas "jamás" lo abandonan. Es bueno hacer
notar que todos estos términos y clasificaciones son relativos; son reglas biológicas
que no son absolutas, ya que nos permiten conceptualizar fenómenos que rara vez
carecen de excepción.
3. Dispersión
Los insectos responden a los cambios ambientales de corto y largo plazo. El cambio más frecuente en su ambiente es la abundancia relativa de alimento; la abundancia de sitio de oviposición o de refugio cambia relativamente menos. Cuando los sitios de oviposición y refugio escasean, o se reduce la capacidad de sustento local, los insectos comienzan a vagar en busca de estos satisfactores. Esto los obliga a disminuir la densidad de población por unidad de superficie, es decir, a dispersarse. Muy frecuentemente, sobre todo entre los no entomólogos que se dedican a la entomología, a este fenómeno lo llaman emigración. Falso; la emigración es parte de un fenómeno periódico y fijo, que no siempre tiene que ver con situaciones coyunturales como el agotamiento temporal de un recurso; además, suele tener la fase contraria o inmigración; ambas configuran la migración. La dispersión normalmente abate la densidad de población local sin modificar las poblaciones absolutas, y se efectúa hacia hospedantes vecinos más susceptibles o simplemente disponibles como alimento, refugio temporal o lugar para ovipositar, mudar, pupar o copular.
Cuando los artrópodos se dispersan para pupar y/o invernar, ya no los consideramos plaga, y lo mismo
sucede cuando se esparcen en busca de hospedantes silvestres locales; esto es
un error porque el manejo ecológico de una plaga implica ser implacable, especialmente
cuando se conocen todas sus bases. Conocerlas bien significa identificar las
posibilidades de supervivencia y las trincheras del artrópodo, que deben ser
conocidas al máximo; sin dejar de considerar los aspectos ecológicos y
económicos.
4. Migración
Son los insectos epidémicos quienes tarde o temprano agotan algún recurso local y tienen que desplazarse localmente en busca de nuevas áreas susceptibles; ya se dijo que esto es una dispersión. Ocurre bajo presiones poblacionales, y “no obedece” a órdenes genéticamente codificadas ex-profeso. Se trata simplemente de competencia intraespecífica, en donde el más apto coloniza otra planta vecina o cercana, antes que sus conespecíficos.
La migración completa es un
fenómeno recurrente que involucra un éxodo o emigración y un regreso a la misma
zona o inmigración. Está genéticamente programada y es ineludible en el ámbito poblacional.
Predomina entre insectos epidémicos, y sería raro entre endémicos (aunque hay
quien sostiene, por ejemplo, que ciertos homópteros equilibristas, huéspedes
permanentes de plantas perennes, están genéticamente programados para emigrar e
inmigrar dentro de su misma planta, según las condiciones ambientales les sean
favorables o no).
Como animales capaces de
"autotransportarse", los insectos cubren todas las posibilidades de distancia.
Se han detectado movimientos con retorno y sin retorno, de unos cuantos metros,
de unos cuantos kilómetros y de cientos o miles de kilómetros.
Las emigraciones sin retorno, de
muchos kilómetros de distancia de la langosta Schistocerca spp son uno de los fenómenos entomológicos mejor
estudiados; técnicamente deben llamarse emigración (porque es genética y sin
retorno al lugar de origen); pero la Schistocerca
gregaria de algunos desiertos de África, viaja al sur en primavera y al
norte en el verano; por lo tanto migra (emigra e inmigra).
Los movimientos migratorios de
las mariposas monarca comenzaron a ser estudiados más tarde porque su impacto
económico es menor, pero los estudios han avanzado mucho en años recientes
debido al ambientalismo que prevalece en los países involucrados; y por
supuesto al interés científico.
Adultos de Cochliomya hominivorax, el gusano de las heridas del ganado o
"gusano barrenador" exhiben movimientos de dispersión que parecen
migraciones, debido a que en el verano van hacia el norte (les “falta frío”) y
en invierno regresan al sur (les “falta calor”), pero para algunos autores no
son verdaderas migraciones ya que no dependen de condiciones extrínsecas e intrínsecas
conjugadas: temperatura, humedad, fotoperiodo, calidad de la alimentación,
cuerpo graso y balance hormonal.